Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Gen Mutasyonu, DNA onarımı ve Transpozisyon
V. ve VIII. Yarıyıllar
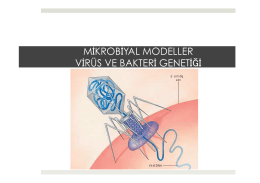
BÖLÜM 18: Mikrobiyal Modeller
TG – 4 - İhtiyaç Yayıncılık
BÖLÜM 5: Makromoleküllerin Yapı ve İşlevleri
Meyve-Sebze Teknolojisi I
6. Bakterilerde ve Bakteriyofajlarda Genetik Analizler ve Haritalama
19. bölüm.pptx - Prof. Dr. Bektaş TEPE
Drosophila - Prof. Dr. Bektaş TEPE
Bilimler Dergi/i - Pozitif Bilimler Dergisi
Murat DURAN - Fen Edebiyat Fakültesi
OSYS Çalıştayı
Kas ve sinir biopsisi
19(1) - Mustafa Kemal Üniversitesi
NOT1
Dokümanı İndirin - doğumsal
Balık 1.1 - Uzman Veteriner
Giriş (Geçmişten günümüze nörobilimler)
barış altay
Ebru Tarım
VİROLOJİ
PC–008 Değişik Biyoinformatik Araçlar Kullanılarak ITS1, ITS2 Gen
Genetik Mühendisliği - Hacettepe Üniversitesi