Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
19. bölüm.pptx - Prof. Dr. Bektaş TEPE
özefagus hastalıkları
P12. Disiplinler Arası Bir Bilim Dalı: Biyoinformatik
Biyoloji 10 - İhtiyaç Yayıncılık
Tam Metin
Kronik Miyeloid Lösemi: Patogenez, Klinik Özellikler
13. Genetik Şifre ve Transkripsiyon.pptx
Veri Şifreleme Teknolojileri
Probiyotik Tedavisinden Sonra Yaşlı Bir Hastada Gelişen
BÖLÜM 12: Kromozomlardaki DNA`nın Organizasyonu
Şifreleme - Securing The Human
Gen Mutasyonu, DNA onarımı ve Transpozisyon
17. BÖLÜM - Prof. Dr. Bektaş TEPE
14. Translasyon ve Proteinler.pptx
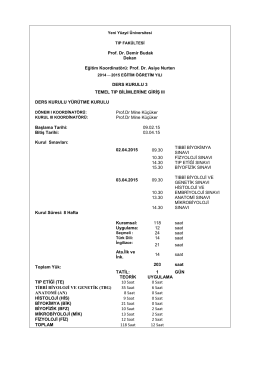
3. Kurul Ders Programı - İstanbul Yeni Yüzyıl Üniversitesi
Ökaryotlarda Gen İfadesinin Düzenlenmesi
DIZEL YAKIT SISTEMLERI ve COMMON RAIL-2015
Aynı Hastada Görülen Üç Farklı Organ Malignitesi
8. bölüm
Lisans Eğitim Öğretim Yönetmeliği (YENİ Türkçe) - IE101
multiGen7L - PDF format
Primer poglavlja
DNA Dizileme ve Filogenetik Analizler