Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
BÖLÜM 7: Ökaryotlarda Kromozom Haritalaması
Kontury vývoje vzdělávání českých novinářů
Nadzorni odbor Fonda zajedničkog ulaganja Trend
Genel Ekonomi - Aktif Akademi Eğitim Merkezi
SORTEC Optical solutions 2014
Drosophila - Prof. Dr. Bektaş TEPE
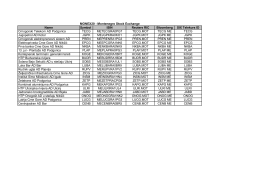
Name Symbol ISIN Reuters RIC Bloomberg SIX Telekurs ID
n-asetil sisteinin endometriyum hücreleri üzerine etkileri
15. BÖLÜM - Prof. Dr. Bektaş TEPE
Mtz-Myz
EKONOMİK TAKVİM GÜNÜN ÖZETİ
Download (486kB) - Munich Personal RePEc Archive
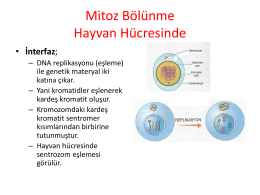
Mitoz Bölünme
Grup 22 - Ankara Üniversitesi
6. Hafta
5. Hafta
2. Mitoz ve Mayoz.pptx - Prof. Dr. Bektaş TEPE
MkaSteel ile - mka yazılım
EBE 238 Ders 2
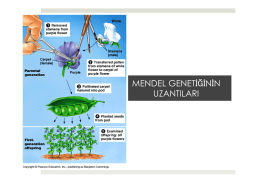
3. Mendel Genetiği.pptx - Prof. Dr. Bektaş TEPE
16. Hafta
2014-2015 10.Sınıf 1.Dönem 1.Notlar
3 Test