Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
2. Mitoz ve Mayoz.pptx - Prof. Dr. Bektaş TEPE
TEK MİDELİLERDE METABOLİZMA
Tarihi II” dersini DÖNEMLİK ALAN ÖĞRENCİLERİN DİKKATİNE!
Fen ve Teknoloji 5 - İhtiyaç Yayıncılık
8.SINIFLAR HAFTA SONU KURS PLANI PAT
Biyoloji-Soru-B
11. BÖLÜM - Prof. Dr. Bektaş TEPE
8. Sınıf - Hazır Bulunuşluk Sınavı
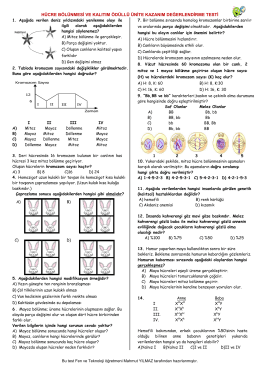
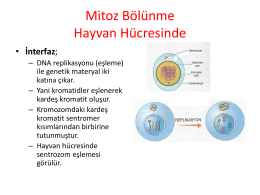
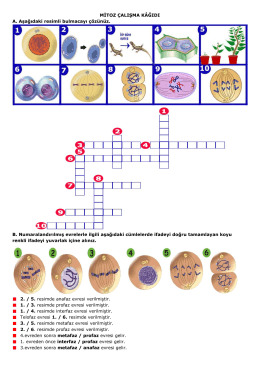
Mitoz Bölünme
6. Bakterilerde ve Bakteriyofajlarda Genetik Analizler ve Haritalama
Mtz-Myz
bölünme - Adnan Menderes Üniversitesi
13. BÖLÜM - Prof. Dr. Bektaş TEPE
İş Jeti Pazarının Geleceği Tahminleri Üzerine... 21 Ekim 2014
Bu dökümanın mobil sürüm için pdf hali
Kanser nedir? - Medikal Onkolog – Prof. Dr. Hüseyin ABALI
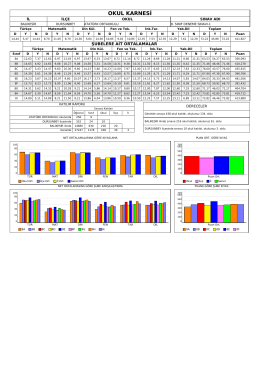
OKUL KARNESİ
8. Sınıf Konu Dağılımları
H - Uludağ Üniversitesi Ziraat Fakültesi
12. BÖLÜM - Prof. Dr. Bektaş TEPE
İndir (PDF, Bilinmeyen) - Biyoloji Okur
Dersimiz Hücre
Canlılığın Temel Birimi Hücre