Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Tam Metin/Full Text - Muş Alparslan Üniversitesi
Işığın Kırılması
2016 yılı insektisit alım işi alımı 4734 sayılı Kamu İhale Kanununun
2.Sayı
prednaska
T-list Lions.cdr
Lökoaraiozis
Untitled - Türk Kardiyoloji Derneği
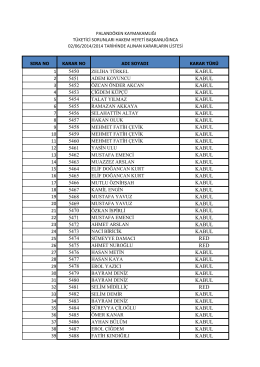
02.06.2014 tarih ve 12 sayılı Tüketici Hakem Heyeti Kararı
Q~ ...lOG 20 i2 - Adana Halk Sağlığı Müdürlüğü
Characterization of a novel xylose isomerase
Rana ridibunda - Biyoloji Kongreleri
Hepatitis C virus genotypes in Adana and Antakya regions
Ziraat Mühendisliği B - Orman Genel Müdürlüğü
Glukoz homeostazisi - mustafaaltinisik.org.uk
Poidła dla bydła i koni
ovde - Hemcof
EVDE BİYOTEKNOLOJİ - Adnan Menderes Üniversitesi
Ekler
Biyolojik Örneklerde İlaç Analizi ECZ 344/8. Ders 22.05.2014 Prof.Dr
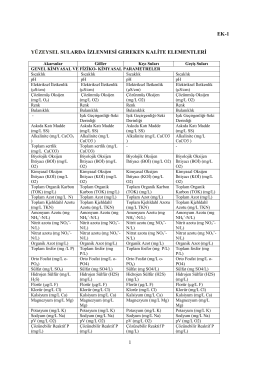
Yüzeysel Sular Ve Yeraltı Sularının İzlenmesine Dair
Vliv selenu na metabolismus laboratorního potkana
Asit ve Asitli Hastaya Yaklaşım - Uludağ Üniversitesi Tıp Fakültesi