Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Sborník
Malárie
Characterization of Internal Transcribed Spacer of
MOLEKULÁRNÍ TAXONOMIE – 9
Parazitologie - úvodní stránky.pdf
Toxoplasma, Pneumocystis, Trichomonas, Leptospira – co mají
Číslo 2
Serum S100B Protein as an Outcome Prediction Tool
Soubor kapitol ze stránek http://www.genetika
CACHEID=d5ff450d-bbc8-4af6-8d10-bde8feb61cc9;Basın Açıklaması-4/b Hak Ediş Kesintisi
(VD.BM_MIK 02_P1 Seznamy vyšetření)
Serie 1 - řešení - Studiumbiologie.cz
Amibiyaz - Türkiye Halk Sağlığı Kurumu
ไวรัสในสิ่งแวดล้อม: ผลกระทบทางสาธารณสุข
institute of vertebrate biology
Spotkanie ze Specjalistą III
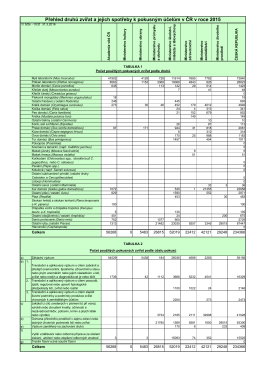
Evidence zvířat použitých v pokusech.
Program Věda na vsi
Obalka zbornik - Česká parazitologická společnost
Wpływ podawania magnezu na zaburzenia metaboliczne u osób z
ZOOREPORT PROFI Nepůvodní druhy savců v České
programem
ZPRÁVA REPORT