Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
İskelet Kaslarının Yapısı ve Biyomekaniği
Uchwała nr
kompletan spisak po nivoima
Miyokardın yapısı
Kas Dokusu
Mišićna jačina
Kas Doku Histolojisi
Besin Allerjileri
SULAMA
9.Sınıf Öğrenci Oryantasyon Sunumunu indirmek için tıklayınız.
P - O nama
azınlık filmleri
CERRAHPAŞA ÖĞRENCİ BİLİMSEL
Aytaç Gökmen
Psödonöbeti olan kadınlarda vestibüler uyarılmıĢ
Zanaatkârlığın Günümüzde Yeniden Yorumlanması: Yeni
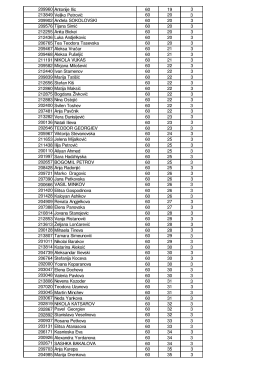
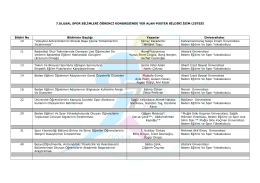
Kabul Edilen Poster sunumlar - 7. Ulusal Spor Bilimleri Öğrenci
IOO 103 SUNU 4
Bitkisel Üretimde Azot Alım Etkinliği ve Reaktif
Determination of elements in dust depositions by
Genel Antrenman Bilgisi
atmosferik geçiş yapan araç etrafında navıer
PowerPoint Sunusu