Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Yerli Sığır Irklarında Myostatin Genine Ait III
âESKÉHO SVAZU CHOVATELŇ MASNÉHO SKOTU
Międzynarodowa Konferencja Naukowa Badania
UÜ-SK BÜTÇE PLAN SATINALMA TEKNİK ŞARTNAME ÖRNEĞİ
Do Rzeczy - 6/2014
20 let chovu Blonde d`Aquitaine „z Rochova“
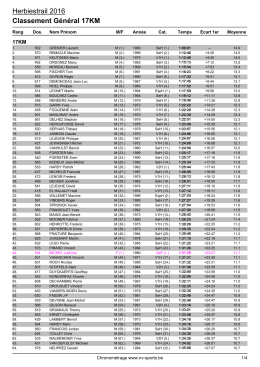
Classement Général 17KM
Paradox telepítői kézikönyv MG 6160-hoz
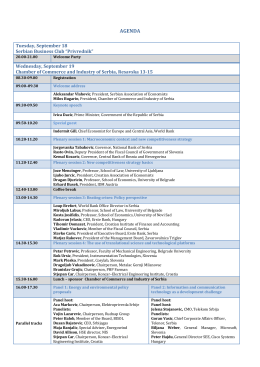
AGENDA
KATALOG - OFFICIAL CATALOGUE
przestępczość farmaceutyczna - konferencja
Full Text - Tarım Bilimleri Dergisi
Plasma cell-free DNA levels in children with Henoch Schönlein
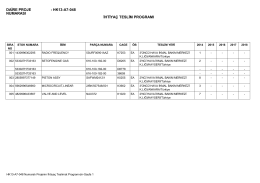
Teslimat programı
kahramanmaraş grubu sporcu katılım listesi
Rekabet İçerisinde İş Birliği”
Diyare tanısında MALDI - TOF MS, Yakut Akyön Yılmaz
1 ağustos 2014 - Ekonomi Gazetesi
Spora bilimsel destek
YOU ARE WHAT YOU BREATHE - BIK
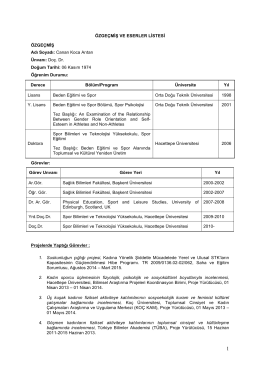
Canan Koca_ozgecmis_2014 - Sporda Toplumsal Araştırmalar
RENCONTRES ECOSSAISES NIMES 2016 conferences
Read Article 297-302 - Turkish Journal of Agricultural and Natural