Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
İmmün Sistemin Tanıtımı
Yaşlılık ve Gastrointestinal Sistem
Doç. Dr. Serdar Ceylaner
Argan yağı
İmmun Yanıt
Üniversitelilerimiz Dûn Seldiler
1 - UniFlip
östrojen ve progesteronun endometr iyum hücrelerinde mitojenle
BÖLÜM 18: Mikrobiyal Modeller
İdrar Sitolojisi Günümüzde Mesane Kanserinin Tanı ve Takibinde
ÇÖLYAK HASTALIĞINDA YENİ GELİŞMELER
Staj listesi için tıklayınız
Prehipertansiyonda epikardiyal yağ dokusu kalınlığı ile kan basıncı
S. aureus - Uzman Veteriner
Makale PDF - Güncel Gastroenteroloji
tustıme nisan 2015 1. tus deneme sınavı
Nörotensin
Kıl follikülü immünolojisi
Hemodiyaliz hastasında anemiye tanısal yaklaşım
Tam Metin (PDF) - Turkish Archives of Otolaryngology
Yrd.Doç.Dr. Ayşe Ceylan HAMAMCIOĞLU
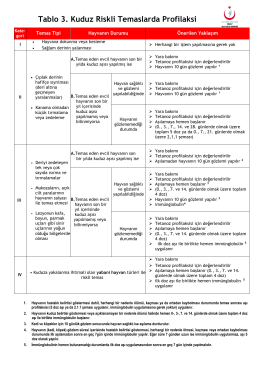
Kuduz Riskli Temas Profilaksisi