Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
9. bölüm.pptx - Prof. Dr. Bektaş TEPE
9. Hücre Solunumu-Kimyasal Enerji Eldesi.pptx
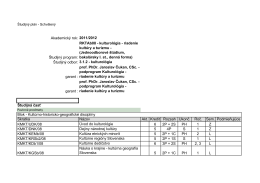
Študijný program Manažment kultúry a turizmu
Sunum için tıklayınız.
DİŞ HEKİMLİĞİNDE UZMANLIK EĞİTİMİ GİRİŞ SINAVI (DUS
AÖF HÜCRE KİMYASI ders notu için tıklayınız!
HİLAL ESEN - Trakya Üniversitesi
11. BÖLÜM - Prof. Dr. Bektaş TEPE
3. Bölüm.pptx - Prof. Dr. Bektaş TEPE
MONO- VE DİSAKKARİD METABOLİZMASI
tusem 2015 nisan 1. deneme sınavı
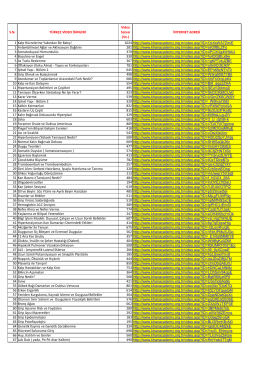
khan academy sağlık vidoları
SOLUNUM
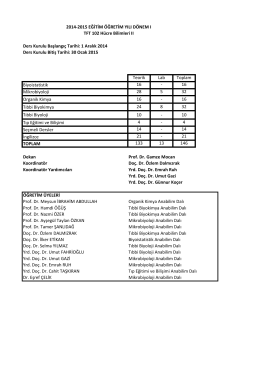
16 Mikrobiyoloji 28 5 32 Organik Kimya 16
YENİ 5,7-DİSÜBSTİTÜE-[1,4]-TİYOAZEPİNLERİN SENTEZİ VE
Ayxmaz/biyoloji Adı: ____4. Güneş enerjisinin
Slnm
Basına ve Kamuoyuna YEŞİL : YOL MU, DOĞA MI, DOLAR MI
Emisja obligacji









![YENİ 5,7-DİSÜBSTİTÜE-[1,4]-TİYOAZEPİNLERİN SENTEZİ VE](http://s2.readgur.com/store/data/000042821_1-4f6e449fbb5ad227504c7eab49df586b-260x520.png)



