Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Bioprospect_1_11.qxd:Layout 1
pdf 1,1 MB
kulturní čtrnáctideník 12. 3. 2014 ročník x advojka.cz cena 39 kč
prospect bi - Biotechnologická společnost
Biologická ochrana proti původcům onemocnění rostlin
4. Vlastnosti vláken
nové úlohy
Úvod do proteomiky
Metody: Stable Isotope Probing (SIP)
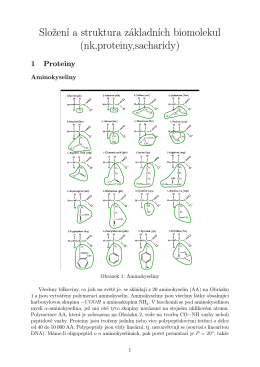
Složení a struktura základních biomolekul (nk
1248 - Ege Üniversitesi Diş Hekimliği Fakültesi
zde
Alergeny z pohledu strávníka
Vznik, projevy a prevence jejich poruch
การศึกษาในปี 2547
Istr. BIONECT Silver Tipo1 Ed.II.ai
Ecka do kapsy.pdf
B I OPR SPECT - Biotechnologická společnost
Present outlook on cardiovascular diseases and statins
Možnosti využití odpadních surovin zpracování vinných hroznů
Vývoj nezaměstnanosti ve Zlínském kraje
Informace o zaměstnanosti ve Zlínském kraji k 30.4.2016
Lipidy II