Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
nové úlohy
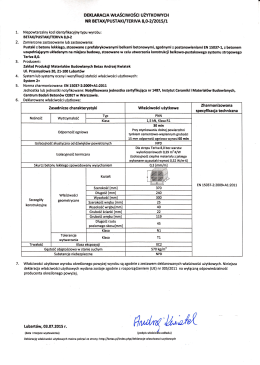
Deklaracja pustak stropowy Teriva 8,0
เทคโนโลยียีนชิป
Špecifikácia prístrojov a zariadení
Bioprospect_1_11.qxd:Layout 1
Výstupy 2015
Skripta - VŠCHT | Ústav Biochemie a Mikrobiologie
Laboratorní příručka oddělení mikrobiologie
Program - Chirurgický deň Kostlivého
3 Pristroje 2013 07.indd
2013–2014 > vybavení pro laboratoře
Univerzita Palackého v Olomouci
ANALIZA PROCESÓW ZACHODZĄCYCH PRZY NORMALNEJ
Diagnostika rostlinných virů - Biologické centrum AV ČR, vvi
Příbalová informace - Abbott Laboratories
Uživatelský manuál
Praha v moderní architektuře pokulhává. Tančící
Úvod do chemie léčiv
sami si zvolte, kolik za telefon u nás zaplatíte títe
MOLEKULÁRNÍ TAXONOMIE - 5 (2014) Protein a
Biofortuna SSPGoTM HLA Wipe Test BF-40
volby 2010 - Nemocnice České Budějovice
Nabídka seminářů pro 4. ročník 2015/2016