Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
bakalarka
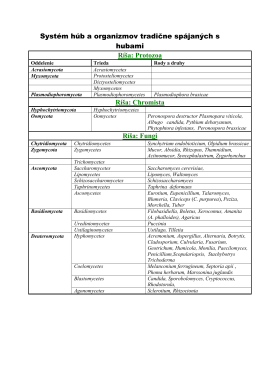
Systém húb a mikroorganizmov tradične spájaných s hubami
Uložit do počítače (PDF, 1.85MB)
Laboratorní příručka - Masarykova nemocnice v Rakovníku
Aeropalynological survey in Büyükorhan, Bursa
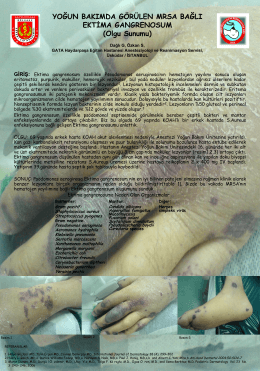
EKTÝMA GANGRENOSUM
Izbornom veću Fizičkog fakulteta Univerziteta u Beogradu
Houby - Otázky z biologie a chemie
157-170 Adem Bicakci.indd - AAI: Astim Allerji Immunoloji
1/2014 Zpravodaj Zdravotního ústavu se sídlem v Ostravě
členové společnosti
Strukoviny
MYKOLÓGIA
Farnosť Sv. Rodiny
Slnečnica a repka
Dodatky k přednáškám
Článek v PDF ke stažení
Požadavky na parametry vnitřního prostředí staveb
Kvalita ovzduší v českých malých sídlech
BEZPEČNOSTNÍ LIST SATUR BADEX
Více v příloze. - oddělení klinické mikrobiologie a autovakcín
Projektowanie i analiza usług sieciowych (PAS)
VYUŽITIE ČISTIČIEK VZDUCHU AIRFREE Ľudia v