Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Магистарска теза
Пробиотиците,
Health Care 6/12 - Комора здравствених установа Србије
Биологија VII тест
Rostoucí mlékárenský průmysl v Číně [PDF]
Zdravstvena nega dece ranog uzrasta I.pdf
Преузмите распоред - Универзитет у Београду Медицински
позив за подношење понуда - Основна Школа СКАДАРЛИЈА
Зборник радова XVI семинара Педијатријске школе
КОНКУРСНA ДОКУМЕНТАЦ ИЈ A Број 1/2015 [Основна школа
Probiotik za živinu – PoultryStar
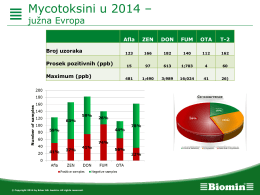
here - ISEO 2014
DON u kukuruzu i svinje
Slabost nogu brojlera i prevencija – Probiotik
Glas Tamnave 127.cdr



![Rostoucí mlékárenský průmysl v Číně [PDF]](http://s2.readgur.com/store/data/000148189_1-e818fabd2e9641a4d6a2b0e2534e3033-260x520.png)