Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
EKOLOGIJA
Osnovi biohemije
ISPITNA PITANJA IZ PREDMETA ŠUMARSKA ENTOMOLOGIJA I
34 UDK 504:631 502:631 UTICAJ POLJOPRIVREDE NA ŽIVOTNU
Sremska poljoprivreda broj 14 26. april 2013.
Преузмите студију затеченог стања
Otvori PDF - Veliki Prasak
NEUROPATIJE UVOD Neuropatije označavaju bolesti perifernog
Zaštita pčela i voska od napada voskovih moljaca i varoe
srbija i klimatske promene
Obrazovni standardi za kraj prvog ciklusa obrazovanja i vaspitanja
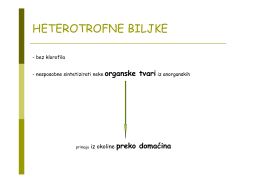
HETEROTROFNE BILJKE
Vitaminlerin İnsan Sağlığındaki Önemi
Визуелна фото хромотерапија
biologija
upravljanje materijalnim i energetskim tokovima