Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
moleküler biyolojide yeni görüşler
ziya gökalp – türkçülüğün esasları pdf
alzheimer-haber-bahar-2015 - Türkiye Alzheimer Derneği
การบําบัดโรคด วย RNA interference (RNAi)
indir - Multipl Skleroz
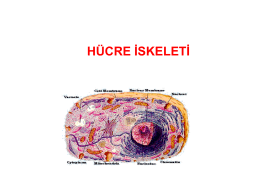
HÜCRE İSKELETİ
lLETl§lM ARA^LARI UZERtNE gALI§MA: KURAMSAL YAKLA
Gelişim Psikolojisi Ek 2
Tam sayfa faks yazdırma
Biologie - přípravný kurz 2011/2012
Arşiv Kaynak Tarama Dergisi
PhD Review Paper / Doktora Çalışması Derleme Makalesi
M492 Hofer HU 2014
Geometrické membránové systémy a modelování cytoskeletu
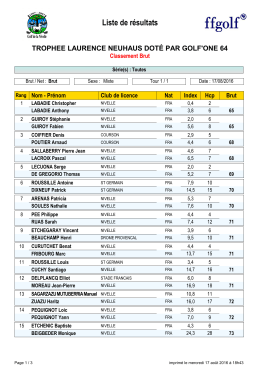
Résultats Bruts - Golf De La Nivelle
Kristal yapı
CURRICULUM VITAE Ireneusz SZCZEŚNIAK September 9, 2015
SLOVENSKÝ - VernostnaKarta.eu
Družstevné Noviny
Norwegia
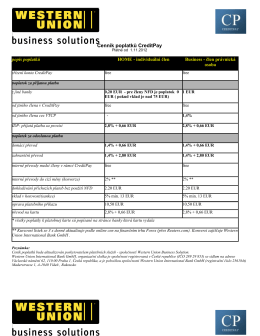
Cenník poplatkov CZ