Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Süt Sığırlarında Postpartum Anöstrüs
ВЕТЕРИНАРСКИ ЖУРНАЛ РЕПУБЛИКЕ СРПСКЕ Veterinary
Wykaz Podręczników 2014-2015
EK – 1 ADANA TİCARET ODASI
Holştayn Düvelerde Sabit Zamanlı Tohumlama Sonrası
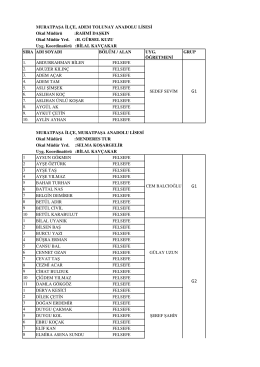
G1 G1 G2
Kalsiyum sabunu ile beslemenin süt verimi, vücut ağırlığı
İneklerde Döl Verimi Sorunları
Weaning
satınalma rehberi - ELSAN Elektrik San. ve Tic. A.Ş.
açılış konuşması metni için tıklayınız
Süt Sığırlarında Gebelik Oranı Üzerine Etkili Bazı Faktörlerin Lojistik
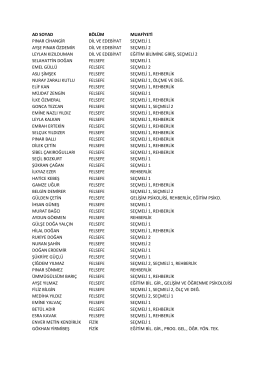
ad soyad bölüm muafiyeti pınar cihangir dil ve edebiyat seçmeli 1
นายเจษฎา จิวากานนท์ - คณะสัตวแพทยศาสตร์
tc yalova üniversitesi sanat ve tasarım fakültesi iletişim sanatları
Enerji İhtiyacının Karşılanmasında Yeni Nesil Yağların
Gonasyl - Luteosyl
yaşlılık ve fiziksel aktivite
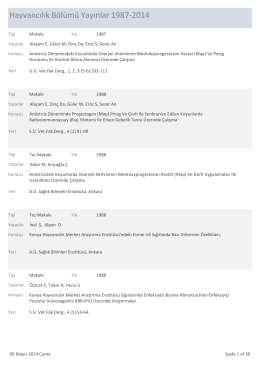
Hayvancılık Araştırmaları Uluslararası Yayınları 1987-2014
Östral ve Luteal Dönemlerdeki Koyunların Oviduktunda Mast
Lax Vox Ses Terapi Tekniği
PROGRAM BIMBINGAN TEKNIS KEPALA LABORATORIUM
Pohlavný cyklus u mačky a psa