Read
Gur
☰
Explore Categories
Sign in
Sign up
Upload
×
Download
No category
The effect of risk elements in soil to nitric oxide metabolism in
bupati badung provinsi bali peraturan bupati badung nomor 21
2014 húsvét - Magyar Katolikus Misszió München
INDUKCE FLUORESCENCE CHLOROFYLU in vivo V PRŮBĚHU V
ovom linku - Tehnika emocionalne slobode
Душица Павловић - Конференција универзитетâ Србије
Bisfosfonátová osteonekroza čelistí.pdf
Univerzita Palackého v Olomouci
AFSA 261
AFSA 302
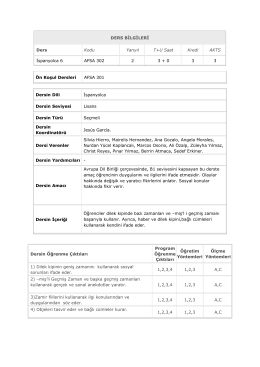
AFSA 301
afsf 202 çeviribilim 4
Doç. Dr. Yeşim Er Öztaş - Hacettepe Üniversitesi Tıp Fakültesi Tıbbi