Read
Gur
☰
Explore Categories
Sign in
Sign up
Upload
×
Download
No category
upper miocene fauna of orešac near smederevo
SZTANÓ O., MAGYAR I., SZÓNOKY M., LANTOS M
XXIII. medzinárodného sympózia Staršia doba bronzová v Čechách
PR Skalní potok - Malacologica Bohemoslovaca
Synthesis 2015 Conference Programme April 16
Preliminary radiocarbon dated paleontological and geological data
Program - eco-ist
dersġn kodu - Ankara Üniversitesi Ziraat Fakültesi
Feuille Paroissiale du mois d`août 2016
Kütüphanemize Yeni Gelen Kitaplar - 2014/1
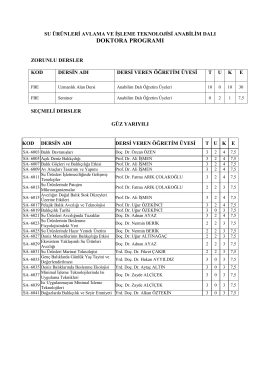
doktora programı
Подкласс Exogenia Collin, 1912
(Hatay) Bulunan Tatlısu Midyelerinin