Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Bu PDF dosyasını indir
Arşiv Kaynak Tarama Dergisi
Bakanlığımıza Bağlı Çanakkale Gökçeada Eğitim ve Dinlenme
ARAŞTIRMA ETİĞİ - Sağlık Bilimleri Enstitüsü
ZDE - Cykloservis
Ebe ve Hemşirelerin Bilimsel Çalışmalara Yönelik Tutumlarının
Obezite ve Deri
6 Kasım 2015 Sayı : 661
GUIDA DELLO STUDENTE - W3.UniRoma1.it
ZDE - Cykloservis
16/03/2015 tarihinde yapılan mübaşirlik mülakatı sonucunda başarılı
HIV pozitif hastada transplantasyon
Dumlupınar Üniversitesi Sosyal Bilimler Dergisi / Dumlupınar
Galaktozemili Bir Yenido¤anda Hipotiroksinemi Hypothyroxinemia
kasım ayı faaliyet raporu için tıklayınız
15.cilt sayı 1-2.indb
MAHMÜTBEY-PAŞADEĞİMMINÎ (İSTANBUL) CtVAM KUtMATAŞ
Umowa grantowa - Nowe Technologie Lokalnie
Zor Entübasyon Düşünülen Olgularda Storz DCI
Newsletter HJF č. I/2012 - Advokátní kancelář Hartmann, Jelínek
PDF İndir - Türkiye Aile Hekimliği Dergisi
Araştırma Makaleleri / Research Articles
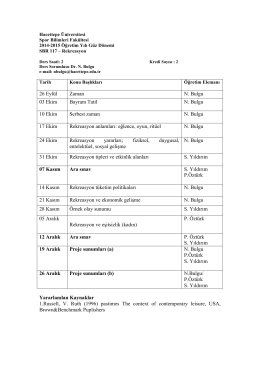
26 Eylül Zaman N. Bulgu 03 Ekim Bayram Tatil N. Bulgu 10 Ekim