Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
KÜLTERE EDĠLMĠġ C6 GLĠOMA HÜCRELERĠNDE
Ovis aries - Biyoloji Kongreleri
6. DOĞAL GAZ (Ref. e_makaleleri, Petrol, Petrol Kimyası, Yakıtlar
RESVERATROL VE EPĠGALLOKATEġĠN KOMBĠNASYONUNUN
IEEE Paper Template in A4 (V1) - Bilecik Şeyh Edebali Üniversitesi
BİTLİS YÖRESİNDE NAR (Punica granatum L
Beyaz Eşya
Manganese-superoxide dismutase (MnSOD) polymorphisms
ÖZET (PDF) - Akuademi.Net
PCOS
ıı. iş fikri yarışması finale kalan projeler
50. Ulusal Psikiyatri Kongresi
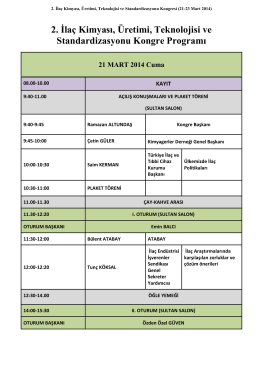
Özet Kitapçığı - 3. İlaç kimyası, Üretimi, Teknolojisi ve
Journal of FisheriesSciences.com
Yrd. Doç. Dr. Ebru HAZAR BODRUMLU
Zelený čaj Matcha: nápoj s výnimočnými vlastnosťami
PA–081 1,4-Dioksan Verilen Albino Farelerde Yeşil Çayın Bazı
Süre ve Sıcaklığın Yeşil Çayda Deme Geçen Kateşinler