Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Employee shift schedule
Laboratuvarda Ergonom Önemldr.
Lista e kandidatëve të subjekteve politike Lista kandidata političkih
Král - Od detekce tumorových markerů k aplikaci 1.část
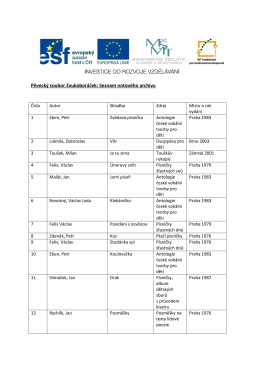
Pěvecký soubor Zvukoboráček: Seznam notového archivu
Izrael je zázrak – jen pro nezasvěcené - Česko
Nedeľa: Tretia „cez rok“
2014-2015 Öğretim Yılı Bitirme Ödevi
Toksidromlar Doç.Dr.Orhan Çınar
Cloning, purification, and characterization of a thermophilic
Full Text PDF - Yüzüncü Yıl Üniversitesi
Yüksek Kretli Üstyapılı Şevli Dalgakıranlarda Hafi f Kret Blokları
Düşen Türk HavaYolları Uçağı nın Enkazı Dün Sabah Bulundu
ANKARA ÜNİVERSİTESİ ZİRAAT FAKÜLTESİ LİSANS
dalgakıran koruma tabakası tipinin, boyutunun ve yerleştirme
สารกําจัดแมลง และศัตรูพืช (Pesticide)
9_em_ ICD 11 geliştirilme.indd
Súťaž – Cena SFaS SLS pre mladého slovenského
fájl megnyitása: semmelweis_irodalom.pdf
3.Ročník Luky
Vážení přátelé historie,