Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
MBG 111 Biyoloji I Fotosentez Aydınlık Evre
VİTAMİNLER
Klientský formát BEST KB (PDF soubor)
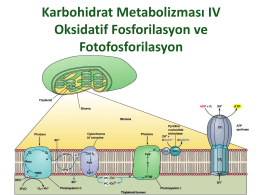
Karbohidrat Metabolizması IV Oksidatif Fosforilasyon ve
2 ATP - Eba
Okuyun - Bilgeler Zirvesi
biyoloji_2013-A - WordPress.com
Tam Metin - Posseible Düşünme Dergisi
H36
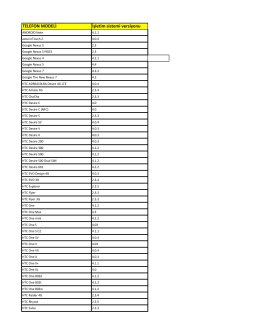
TELEFON MODELİ İşletim sistemi versiyonu
2012_ulusal_biyoloji_A_kitapcik
kimya bilimi - WordPress.com
element kavramı
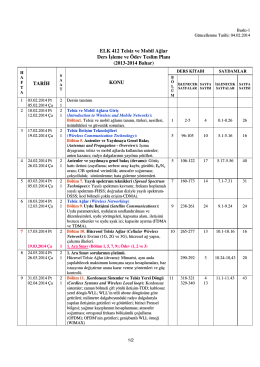
ELK 412 Telsiz ve Mobil Ağlar Ders İşleme ve Ödev Teslim Planı
Robot Bilimi Robot Algılayıcıları
yaprak - FaPGenT
Elektromanyetİk Dalgalar
her şey geçip gider
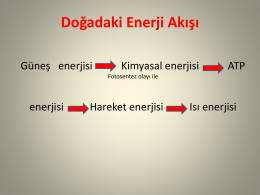
IŞIK ENERJİSİ ve HAYAT
TBBE2303 Biokémia II
İndir - Elazığ Belediyesi
00_Jenerik 23.indd
Deniz Coskun - İstanbul Kültür Üniversitesi