Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Karbohidrat Metabolizması IV Oksidatif Fosforilasyon ve
MBG 111 Biyoloji I Fotosentez Aydınlık Evre
biyoloji_2013-A - WordPress.com
Amino Acid Degradation
Populasyon Ekolojisi - Sakarya Üniversitesi
Hücre ve Hücre Organelleri
Biyoloji 8 - İhtiyaç Yayıncılık
Fotoğrafın 200 Yıllık Serüveni
TG – 2 - İhtiyaç Yayıncılık
Ders13_Fosfolipidler - Prof. Dr. Suat Erdoğan
2 ATP - Eba
Hafta - 10 (Sunum-9)
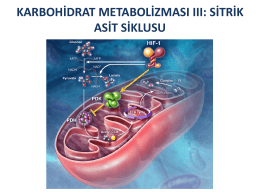
KARBOHİDRAT METABOLİZMASI III: SİTRİK ASİT SİKLUSU
populasyon ekolojisi
FARKLI OZON JENERATÖRLERİ İLE FARKLI
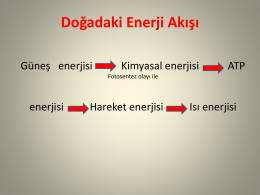
IŞIK ENERJİSİ ve HAYAT
Yeni Microsoft Office Word Belgesi _2
Ayxmaz/biyoloji Adı: ____4. Güneş enerjisinin
SERamik
onlıne katalog
sınav örnekleri
ya da H
cihangir altunkıran