Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
00 yeni.indd - Türk Hematoloji Derneği
Pacient Základní diagnóza Rodné číslo Ostatní
Suçiçeği aşısı sonrası gelişen Ramsay Hunt
Olgu MA - Klimik
Hlas sv. Cyrila a Metoda - Podzim 2013
Susurluk Rapor Kış - Çevre ve Şehircilik Bakanlığı
č.4 - Maskil
Gymnázium Václava Hraběte, Hořovice, Jiráskova
OPIS TECHNICZNY DO CZĘŚCI KONSTRUKCYJNEJ
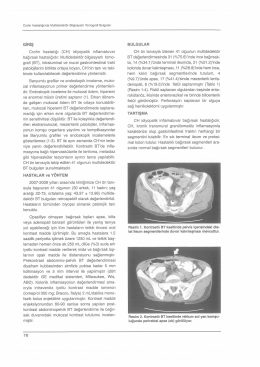
GİRİŞ Crohn hastalığı (CH) idiyopatik inflamatuvar
Transplantasyon sonrası immünolojik monitorizasyon ve önemi
erişkin orak hücre hastaları için transplant protokolü geliştirme
นส. จริยธรรมการวิจัยที่เกี่ยวข้องกับมนุษย์ อ.นิตร มรกต
689-706 Fatih Sahiner.indd
1 ÖZGEÇMİŞ 1. Adı Soyadı: Hasan Fehmi Ateş 2
Lomber Disk Herniasyonunda Neovaskülarizasyonla
Malba: Eva Wimmerová, 12 let
anestezi sınıfı pdf indir - Kariyer Koleji`ne Hoşgeldiniz
Zpravodaj Svobody zvířat
Metakronöz Timoma ve Küçük Hücreli Dışı Akciğer
BRinfo 09-09-2014
7. hét
Alph. Eigentümerverzeichnis der Stadt Tolkemit, Kr.Elbing Tolkemit