Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
İĞNECİKLİ SİNİR AĞLARI
Biyoloji Öğretmenliği TG_9
19. Ulusal Sıvıhal Sempozyumu ( 19 Aralık 2015)
Yapay Sinir Aglari
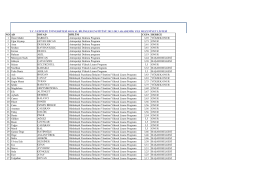
NO AD SOYAD BÖLÜM CGPA DERECE 1 Ömer Mahir SARIGÜL
Eki için tıklayınız
Aleksitimi: Tedavi Girişimleri-Alexithymia: Treatment Interventions
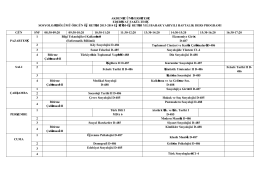
Program - Süleyman Demirel Üniversitesi
Sürekli 2 Bant Boyama ve Apre Makinesi MFR SE 1
kaba daneli zeminlerin sükunetteki toprak basıncı
İÇİNDEKİLER / CONTENTS
Asst. Prof. Dr. Ergin YILMAZ
Dünya Bankası Küresel Ekonomik Beklentiler Raporu
akdeniz üniversitesi edebiyat fakültesi sosyoloji bölümü örgün
PhD Review Paper / Doktora Çalışması Derleme Makalesi
Brožúra na stiahnutie
Yrd. Doç. Dr. Ergin YILMAZ - Bülent Ecevit Üniversitesi Fizik Bölümü
Örnek karar metni
GL544 A/B - GL744 A/B
Ders Notu
GL525 F/H-GL625 F/H-725 F/H "SbyS"
Program ke stažení ()
DE 60 - SE 60 Köşe Takozu