Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
kap. 3
Nasza szkoła - Nasz Elementarz - Ministerstwo Edukacji Narodowej
Základní barvy konstrukce
część 4 - Nasz Elementarz - Ministerstwo Edukacji Narodowej
Natalia MORGULEC-ADAMOWICZ
ภาคธุรกิจที่ไม่ใช่สถาบันการเงิน
vektora a inzertu. Tieto techniky sa preto oznacuj& niekedy aj ako
kunthi - Kinige
()&0
[붙임2] 1605_대학생 집단연수 운영 안전 확보 매뉴얼(보완)
การจัดการเชิงกลยุทธ์ บริษัท สยาม พี.ซี.เอส. จาก
Ç_Ü_TIP FAKÜLTESİ AKADEMİK PERSONEL LİSTESİ
ESKİŞEHİR CİVARININ JEOLOJİSİ VE SICAK SU KAYNAKLARI Bu
Bulletin 2/2012 - Česká společnost pro mechaniku
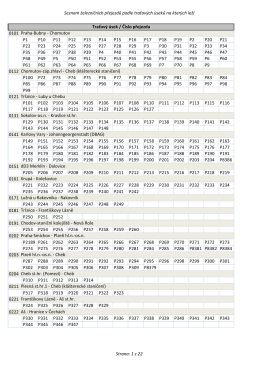
Seznam železničních přejezdů podle traťových úseků na kterých leží
Hereticus 2004/2
абгдж identifikсciu a pre6istenie fragmentov DNA. Medzi jej vhhody
2014-2015 Eğitim-Öğretim Yılı Yaz Okulu
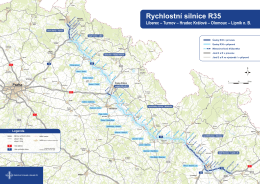
Rychlostní silnice R35
π ·i - Ichel Express
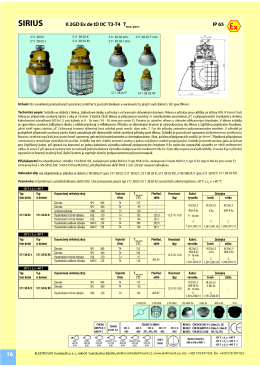
SIRIUS
ç - Ichel Express
z dnia 11 marca 2016 r.







![[붙임2] 1605_대학생 집단연수 운영 안전 확보 매뉴얼(보완)](http://s2.readgur.com/store/data/001656923_1-9adcfc856ec94fef0425748294af5557-260x520.png)