Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Cancer chemopreventive effect of dietary Zataria multiflora
“Ders çalış!” diyorlar ama kimse nasıl çalışacağımı söylemiyor
어린 가축과 초유와 우유
Genç erişkinlerde dumansız tütün “Maraş otu”
Bülteni - Turkish Cargo
Antidot_Listesi
Anoxybacillus sp. 13K Bakterisinden Termofilikrabinofuranosidaz
Preparation and spectral properties of products of the reaction of N
Irodalmi adatok alapján
akademik öz-yeterlik - Eğitim ve Öğretim Araştırmaları Dergisi
10.10.2014
Oya ERYİĞİT GÜNLER - KMÜ Sosyal ve Ekonomik Araştırmalar
i FARKLI RENKTEKİ ALIÇ MEYVELERİNİN POMOLOJİK VE
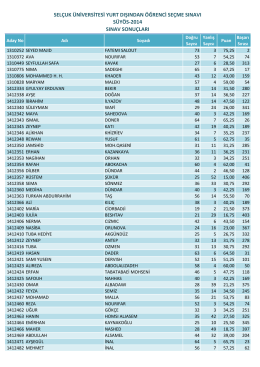
selçuk üniversitesi yurt dışından öğrenci seçme sınavı süyös
Toplumsal Pazarlama Kavramı
ไวรัสร้ายของลูกน้อย…โรคอาร์เอสวี (RSV)
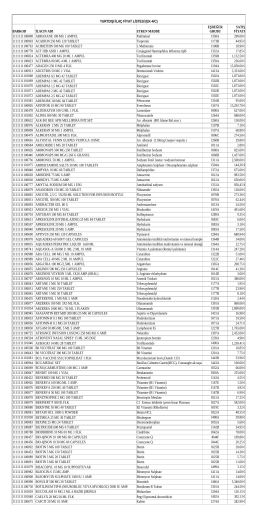
YURTDIŞI İLAÇ FİYAT LİSTESİ (EK
ISSN 1300-0861 E-ISSN 1308-2817 - Ankara Üniversitesi Dergiler
การประยุกต์ทฤษฎีระบบเกรย์เพื่อประเมินการปฏ
Biyomarker Olarak Sitokrom P450 Ekspresyonunun Değerlendirilmesi
HTML – osnove
İstanbul Üniversitesi Eczacılık Fakültesi Biokimya Anabilim Dalı
Real-time PCR analysis of pyrethroid resistance in Helicoverpa