Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Food caching by a Eurasian otter
การระบุสายพันธุ์หน้าวัวโดยใช้ดีเอ็นเอบาร์โ
1 Cvicen´ı z predmetu KMA/PST2 Pro zıskánı zápoctu je
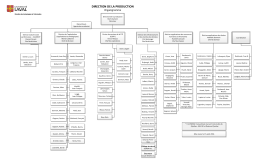
direction de la production - Direction des technologies de l`information
ปุย
Vydra riečna
Sesja plakatowa Kardiologii Zachowawczej i Dziecięcej
Comparison of track and direct observation estimations for
Untitled - ALKA Wildlife
4AM/rtfolio_EN 01 – 07 / 2012
Meris brošura
Comparison of the efficacy of prednisolone, montelukast
Výroční zpráva 2013 - Ústav biologie obratlovců, vvi
Eğitim Kurumlarına Yönetici Seçmede Bir Karar Destek Sistemi
PDF
Studija izvodljivosti za otvaranje državnog
null
Suivez le guide, un voyage au-delà de l`imagination
การออกแบบและก่อสร้างเขื่อนขนาดใหญ่
2 890 51 m2 - gatahouse.pl
Aspekty naukowe związane ze szkodami w gospodarce rybackiej