Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Studium polymorfismu u vybraných populací
VLÁDA ČESKÉ REPUBLIKY PROGRAM SCHŮZE VLÁDY ČESKÉ
Na‰e skúsenosti s vy‰etrovaním JAK2 mutácií pacientov s
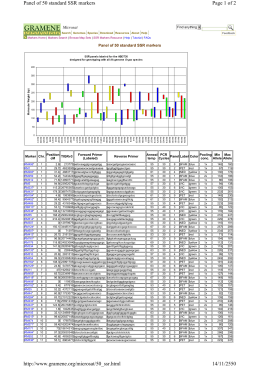
Page 1 of 2 Panel of 50 standard SSR markers 14/11/2550 http
Wykład 3
Dynamika lesa a krajiny jako podklad pro zonaci NP, aneb co
Návod k použití - BAG Health Care GmbH
Appel à projets AMIF 2014-2020 partie générale
zde - Botanický ústav AV ČR
Témata
zde
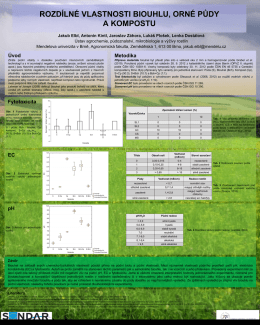
Rozdílné vlastnosti biouhlu, orné půdy a kompostu a
PDF datoteka
Ochrana na úrovni populací: od lokální populace k metapopulacím
A teljes cikk letöltése PDF-formátumban.
Stáhnout soubor - Česká zemědělská univerzita v Praze
Ochrana přírody z pohledu biologa
Artemis Katalog 2015 SON HALI
MOLEKULÁRNÍ METODY V EKOLOGII MIKROORGANIZMŮ
Genetická analýza starokladrubských koní pomocí
Analizy populacyjne. Genetyka w archeologii i kryminalistyce
(Paykull, 1798) (Coleoptera: Dytiscidae) tatranských
Nudmamud-Thanoi S - Naresuan University