Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Research news
fatih sultan mehmet vakıf üniversitesi llp
Yazı için tıklayınız. - Alaşehir İlçe Milli Eğitim Müdürlüğü
TECT 3HAfbA
Dikkat Eksikliği ve Hiperaktivite Bozukluğu Olan Çocukların Anne
Večni mir. Filozofski nacrt - Istorija političkih teorija
NÁZEV PROJEKTU ČÍSLO PROJEKTU JMÉNO HLAVNÍHO
Warszawa, 4 - Mazowiecka Chorągiew Harcerek ZHR
! " $ (* 6 8"
Book of Abstracts - Pannonian Plant Biotechnology Association
Detection of kinase amplifications in gastric
วิชา ชีววิทยา - ทรูปลูกปัญญา
ÖZET (PDF) - Akuademi.Net
Přehled nabídky témat stáží studentů SŠ na VŠ a ústavech
Protokół kominiarski (typ pliku: .pdf ,rozmiar: 1493.67 kB)
fatih sultan mehmet vakıf üniversitesi llp
M. Lenartowicz,Ł.Łukasiewicz
Program 35. semináře ENERGOCHEMIE
Warszawa, 4 - Mazowiecka Chorągiew Harcerek ZHR
Pročitaj više - Veleučilište Baltazar Zaprešić
Výmenné koncovky rámov Author 2001 - 2013 a AGang 2010
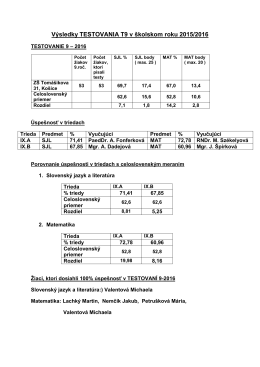
Testovania 9 2016