Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Site-directed mutagenesis of cephalosporin C acylase and
Akademia Ekonomiczna w Katowicach
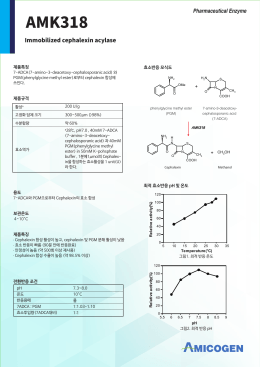
국문 AMK318 Cephalexin
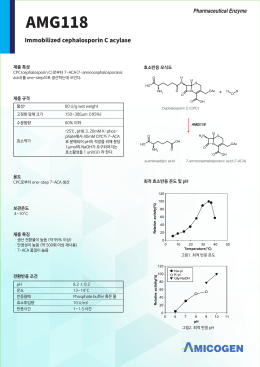
국문 AMG118
06331076 190 100 Elecsys 2010 MODULAR ANALYTICS E170
LML-710 (LMLT-711 Vysielač + LMLR-710 Prijímač
térk termékismertet és árlista - SW Umwelttechnik Magyarország
shmyo_butunleme_2014_bahar
Anaphylactic Shock Developing in Association with Cefuroxime
Sefaleksin Kullanımına Sekonder Gelişen Purpurik İlaç
Ústředny EPS Ústředny řízení SHZ