Read
Gur
☰
Explore Categories
Sign in
Sign up
Upload
×
Download
No category
Arşiv Kaynak Tarama Dergisi
T.C. GAZİOSMANPAŞA ÜNİVERSİTESİ
PPKE ITK - Oktatói publikációk 2012
SFB 15 - Lidl Service Website
SBM 52 - Lidl Service Website
www.febs-lysosome.org
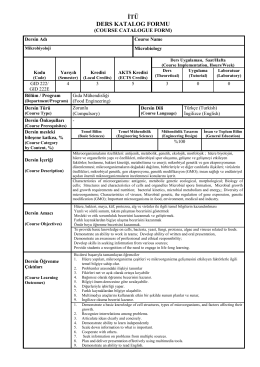
GID 222 - İTÜ Gıda Mühendisliği Bölümü
Tam Metin (PDF) - Turkish Archives of Otolaryngology
Elif Damla ARISAN İletişim Bilgileri
Statik Egzersizin Beyin Hemodinamisine Etkisi
Süte uygulana ön işlemler
NANOTECHNOLOGY AND TISSUE ENGINEERING: CURRENT
Effect of colchicine on experimental acetic acid induced colitis