Read
Gur
☰
Explore Categories
Sign in
Sign up
Upload
×
Download
No category
Tam Makale/PDF - İ.Ü. Tıp Fakültesi Dergisi
vybrané lingvistické aspekty akademického a odborného anglického
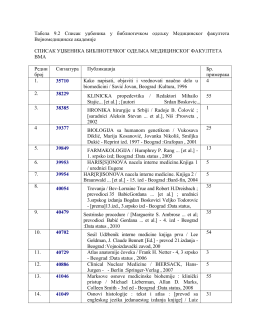
Табела 9.2 Списак уџбеника у библиотечком одељку
maGazíN - expert Elektro
Podstawy zarządzania
Mersin Adliyesi Sözleşmeli Mübaşir Sınavı Nihai Başarı Listesi
POPIS TEMA, MENTORA, ČLANOVA KOMISIJE I OSNOVNE
Table 1 - DergiPark
zootaxa - Magnolia Press
Letöltés PDF formátumban
Oral insülin - Türk Diabet Cemiyeti
T23_b: „Handout” of the lecture
okumak için tıklayın