Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Cloning, purification, and characterization of a thermophilic
program kitapçığı - ekoloji 2015 vı ulusal ekoloji sempozyumu
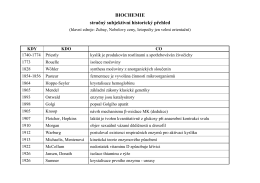
Historie biochemie
Characterization of a novel xylose isomerase
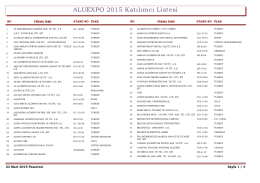
Katılımcı Listesi
Enzimlerin Genel Özellikleri, Sınıflandırılması ve
Anoxybacillus sp. 13K Bakterisinden Termofilikrabinofuranosidaz
yıldızlar ferdi sonuçlar - İstanbul İl Yüzme Temsilciliği
Cloning and expression analysis of 1-deoxy-D
ATPaz Özelliğine Sahip Olduğu VarsayılanAlüminyuma Dirençlilik
Employee shift schedule
İnguinal kanal
สารประกอบใดที่ไม่ใช่น้ำตาล
Optimization of the xylanase production with the newly isolated
Enzim Kine ği (Hızı)
PQ process by WHO
Birleştirme Kaynağı Kaynak Sarf Malzemeleri Çok Satanlar