Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Celiac Disease and Autoimmune Thyroid Diseases
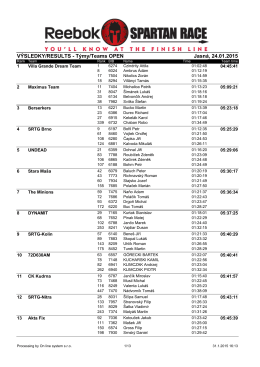
VÝSLEDKY/RESULTS - Týmy/Teams OPEN Jasná
Arşiv Kaynak Tarama Dergisi
Celiac Disease and Autoimmune Thyroid Diseases
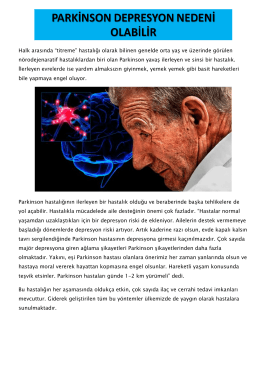
1. Parkinson Depresyon Nedeni Olabilir - Bey-Om-Sin
Celiac Disease and Autoimmune Thyroid Diseases
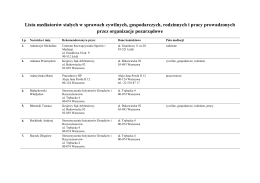
Listy mediatorów stałych w sprawach cywilnych, gospodarczych
Celiac Disease and Autoimmune Thyroid Diseases
Program
Celiac Disease and Autoimmune Thyroid Diseases
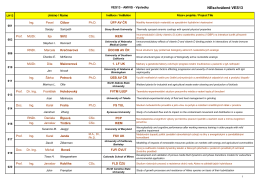
VES13 - AMVIS
16. Ukladanie železa v bazálnych gangliách
Benign prostat hiperplazisi ile ilişkili alt üriner sistem
ZDROJE ENERGIE - Klub zdravia Walmark
„A fürdővezető szakmérnök szakirányú továbbképzési szak képzési
CENTRUM PRÁVNÍ KOMPARATISTIKY
Sağlık Bakanlığı Ordu Üniversitesi Eğitim ve Araştırma Hastanesi
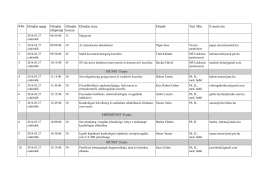
Ewidencja mediatorów stałych w sprawach cywilnych i gospodarczych
Celiac Disease and Autoimmune Thyroid Diseases
Celiac Disease and Autoimmune Thyroid Diseases
RENCONTRE TOUS CHERCHEURS PARC DE LUMINY AIX
23 EKİM 2014 - Ekonomi Gazetesi
Lojistik Sektöründe Sürdürülebilirlik Uygulamaları