Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
DD2008.pdf - Arborétum Mlyňany SAV
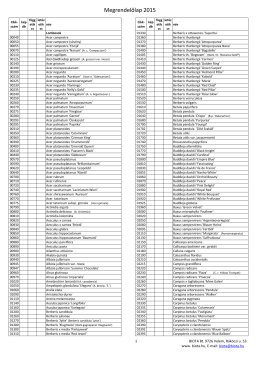
Megrendelőlap 2014
Slovenská akadémia vied
ประเทศและเมืองหลวง
FAKULTA EKOLÓGIE A ENVIRONMENTALISTIKY TECHNICKÁ
Hlucháň a jeho les - tetrao carpathians
Acta 99 - Výzkumný ústav Silva Taroucy pro krajinu a okrasné
priručnik za integralnu proizvodnju i zaštitu crnog luka
Süt Ürünlerinde Tüketici Tercihini Etkileyen Faktörler ve Gıda
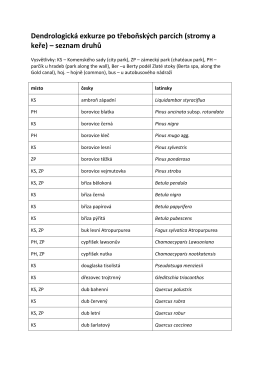
seznam dřevin - stále se aktualizuje
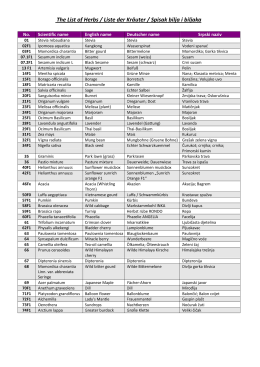
The List of Herbs / Liste der Kräuter / Spisak bilja i biljaka
AMZO-Zahradni_tvorba.pdf 81KB Jan 16 2014 09:47:17 AM
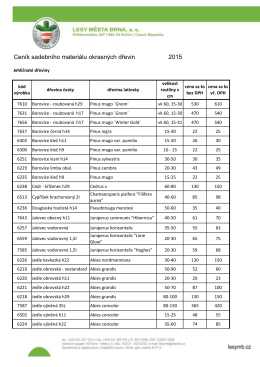
Ceník sadebního materiálu okrasných dřevin 2015
Newsletter 9
278/2012 Z.z. VYHLÁŠKA Úradu pre reguláciu sieťových
1/2012 - Fakulta prevádzky a ekonomiky dopravy a spojov
Przedmiar.pdf
C. Komplexná charakteristika a hodnotenie vplyvov na životné
of oak galls from inner western Anatolia, Turkey
Variability in the growing season of selected European and East