Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
elektroakustický prístroj na určovanie úrovne
ZDRAVOVEDA PRE STREDNÉ ŠKOLY
Zde
Latanoprostum 50 μg + Timololi maleas 6,83 mg
Velkonocny receptar
Osobná hygiena
1. ročník
Vypracované otázky RMA
แนวทางปฏิบัติในการให้บริการทางกิจกรรมบําบั - กรมการแพทย์
12 prednaska
Slovenský variant vyvážacej súpravy naplno boduje!
36. Fázová analýza pomocou Mössbauerovej spektroskopie
1304340734_ziarive_v..
Výročná správa za rok 2013 - HARMÓNIA
3.LINEÁRNE ANTÉNY Lineárnymi anténami rozumieme také antény
väzba – [l - datasolution.sk
2011/3 - modarevue.sk
aciklovir – inhibítor DNA-polymerázy, drivát
doprovodný program kongresu Pražské dentální dny
viscerocranium →viscerokránium
ZDROJE NEUTRÓNOV
Ispit, januarski 2 rok, 17.03.2012
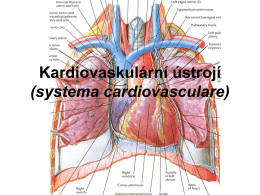
anatomie_kardio.pdf9.76 MB