Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Real-time PCR analysis of pyrethroid resistance in Helicoverpa
oz_bredova_kovacova.pdf
Işığın Kırılması
244 - Türk Biyokimya Dergisi
31-Sprawozdanie OTK 2015_4
Products
tc balıkesir üniversitesi fen bilimleri enstitüsü kimya anabilim dalı
Cancer chemopreventive effect of dietary Zataria multiflora
POLICJA WARMIŃSKO-MAZURSKA DOBRE MIASTO: SPOTKANIE
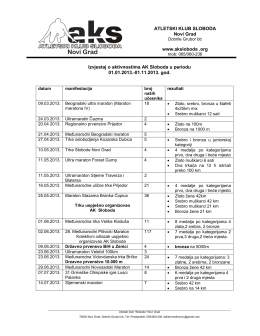
ATLETSKI KLUB SLOBODA Novi Grad www.aksloboda .org Izvjestaj
1 Na osnovu člana 54. i 139. Statuta Univerziteta u
„Planuj karierę i pracuj bezpiecznie” Ogólnopolski Tydzień Kariery
Proje ve Makaleler
ความต้านทานของหนอนเจาะสมอฝ้ายที่มีต่อ d
Hepatitis C virus genotypes in Adana and Antakya regions
Proje ve Makaleler - Mehmet Serdar Güzel
Zawsze proś o oryginalne części zamienne Feraboli - San
u .pdf dokumentu