Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Vibrace BCM094.pdf
β-CrossLaps/serum - Roche Diagnostics
Lista 1 (Wprowadzenie)
null
Karta pracy
27 NMR VALTICE
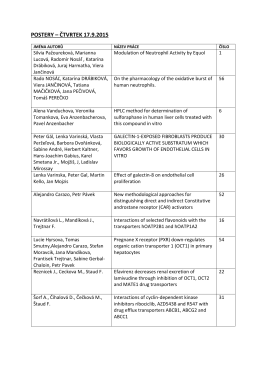
POSTERY – ČTVRTEK 17.9.2015
Pobierz
Phyteneo Žaludeční kapky 20 ml PHARMACEUTICALS
dosyayı indir - muharremyavuz
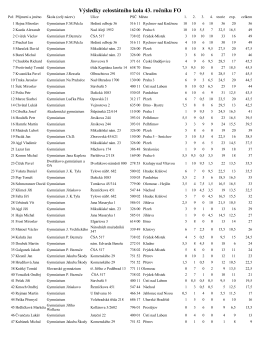
Výsledky celostátního kola 43. ročníku FO
ulusal pıyano yarışması - Hacettepe Üniversitesi
PRO ZDRAVÉ TĚLO ABM (Agaricus Blazei Murrill
Genel Kimya - 15 - Organik Kimya
Úvod do chemie léčiv
Yangın - Abdurrahman İNCE
Výběr z aplikací Ramanské spektrometrie
TITLE OF THE PAPER (TNR 12p size, bold style, upper case, left
věda a výzkum - Akademie věd ČR
univerzita komenského v bratislave prirodovedecká fakulta využitie
Hodnocení stavu vývrtu hlavně s využitím
spracovali zvláštny dokument
CV İçin Tıklayınız.