Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Úvod do molekulární biologie
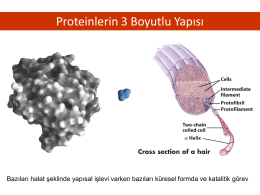
Proteinlerin 3 Boyutlu Yapısı
1 - Biochemie
Ohroženost vodní erozí
Metody: Stable Isotope Probing (SIP)
Reologie - aplikace.pdf
Otázky
Výuka chemie pro nechemické obory na vysokých
Jak (ne)funguje imunitní systém
Obecná zoologie - KATEDRA BIOLOGIE
Zdraví národa Autizmus u detí (SK) Sexuálně
laboratorní postupy při zjišťování půdních vlastností – analýzy
rozhovor k projektu LONGWOOD
Oddělení medicinální chemie– Organická chemie
zde - cykloseminare.cz
ve formátu PDF
Fosfor - maturitní otázka z chemie - Biologie
ČVUT - Fakulta jaderná a fyzikálně inženýrská Katedra jaderné
Bez nadpisu - Slovenská poľnohospodárska univerzita v Nitre
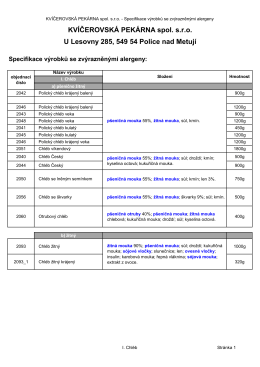
Pekařské alergeny - Kvíčerovská pekárna
Složení nebalených výrobků a minimální trvanlivost