Read
Gur
☰
Explore Categories
Sign in
Sign up
Upload
×
Download
No category
MYKOLOGIE
Aridní oblasti
สาหร่ายสีเขียว - คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล
Abstracts - CZECH MYCOLOGY
zázrak života - Scientific American
Biodiverzita cévnatých rostlin, lišejníků, sinic a
mykorrhizní symbióza
Ascomycota – houby vřeckovýtrusé
Nebuněční – Viry, viroidy, priony
2.cvičení studijní materiály a pracovní listy
ZDE
pobierz
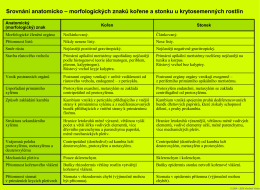
Srovnání anatomicko-morfologických znaků kořene a stonku