Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
vypracované otázky z fyziologie 2009/2010
Otázky
Tvrdé souhlásky
Obecná zoologie - KATEDRA BIOLOGIE
kapitoly z biologie člověka - Další vzdělávání pedagogických
Stáhnout v PDF - Biologická olympiáda
a neuroanatómia vizsga követelményei fogorvostanhallgatók számára
Lázeňské listy - Luhacovice.cz
Vypracované otázky z mikrobiologie
Nemocní s průlomovou bolestí si umějí vážit života
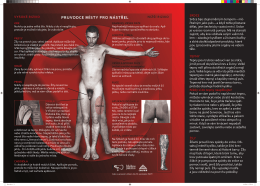
Průvodce místy pro nástřel
„Pitný režim“ a „otrava vodou“
ke stažení
2012 - Fakulta životního prostředí - Česká zemědělská univerzita v
Kapitola 9 Nemoci ledvin, skripta
I rostliny mají hormony [pdf]
Skripta Základy neurověd
I m u n o l o g i e 98 - Centrum klinické imunologie
Découvrez... La Namibie
Selhávání ledvin
Stefano Erkolino
tímto odkazem












![I rostliny mají hormony [pdf]](http://s2.readgur.com/store/data/000152782_1-44a40b4f530386dabc9821997c460986-260x520.png)





