Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Sphaerechinus granularis
Linkster sedlá sedlovky upínaky.pdf
TEST PITANJA IZ BIOLOGIJE
eray çalışkan -embriyo seçiminde array cgh
LİTYUM Yeşin Esin, Lütfiye Eroğlu İstanbul Tıp Fakültesi Farmakoloji
PhD Danijela Joksimovic CV
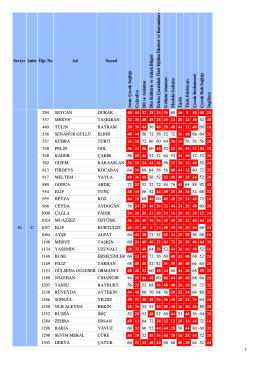
10/C
Preuzmite broj 38.
SİNİR SİSTEMİ HİSTOLOJİSİ
Pobierz Menu - Mazurski Raj
Jeseníky
Početak života
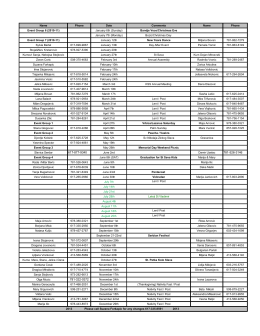
January 6th (Sunday) - St Sava Serbian Orthodox Church
Medicina zasnovana na dokazima
Francouzská pošta testuje elektromobil, jehož dojezd
CFA COMPACT FAN AIR SFA STANDARD FAN AIR
vrozené vývojové vady
N°629-A Notre Eglise 2016-07-31
History of Bible Translation
instrukcja instalowania i obsługi
Télécharger le dossier de presse pour en savoir plus
Istrazivacki rad u masinstvu, Metod NIR
Document