Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
29. Ľudské leukemické bunky sú schopné výrazne
Prenosové Systémy
Otevřít - Onkologie
Non-Steroid Anti-İnflamatuar İlaçlar (NSAİİ)
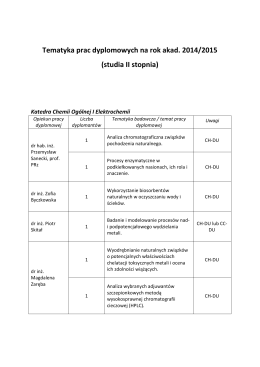
studia II stopnia
mladí vedci – bezpečnosť potravinového reťazca
Čo sa chystá budúci rok - Slovenský spolok študentov farmácie
Financované projekty VEGA z ústavov SAV v roku 2012
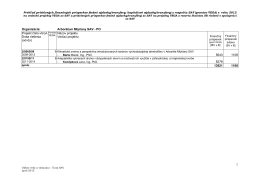
Registr AML-BFM 2012 Versions-Nr1, 1
Identifikácia prekurzorových CD34+ buniek v regenerujúcej
Analógovo-číslicový prevod I.
Emulsje wielokrotne jako mikrośrodowisko do badań biologii komórki