Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Aktuálne poznatky o hliníku v úlohe vakcínového adjuvantu
Vakcíny: Tu je úplný príbeh
Opinia do strategi walki z rakiem
Ako môže by Ako môže byť Ako môže byť niečo tak
Hliník je tretím najrozšírenejším prvkom v zemskej kôre a je prítomný
15 1. V prípade výskytu jednoročných tráv alebo pýru je možné
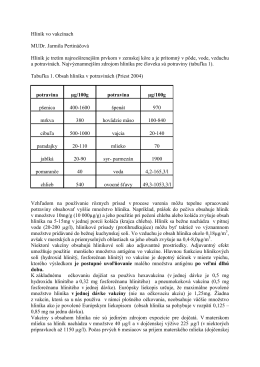
Hliník vo vakcínach
Konspirace versus vědecká pravda?
Aktuálne poznatky o hliníku v úlohe vakcínového adjuvantu
Herbicyd Chikara 25 WG
Hliníkové nanočastice v mozgu
rozhodnutie o odvolaní - stavba Oplotenie v obci
VV P 6-01 - Štatistický úrad SR
la démarche expérimentale
19 - RizikaOckovania.sk
RNDr. Marek Petráš Komentář ke článku Imunologie hlinitých solí ve
ZMLUVA O DIELO číslo: 0606/20130 Objednávateľ
İndir (Türkçe PDF)
Kolektívna imunita - mýty a fakty
Bábätko zomrelo po podaní 9 vakcín v jeden deň
KNIHY O OČKOVANÍ - Sloboda v očkovaní
Strunecka-odpoved-doc-Madarovi
Osýpky, mumps a rubeola - MMR