Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Likelihood reinstates Archaeopteryx as a primitive bird
P t á c i (Aves) I.
Dzieje 7
madarea2014_elso [Kompatibilitási mód].pdf
หุบเขาไดโนเสาร์ในทะเลทราย Gobi
wniosek o przyjęcie dziecka do świetlicy
Opis przedmiotu zamówienia
ไม้หินฯ จัดสัมมนา ณ จังหวัดสมุทรสงคราม
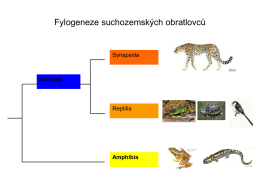
21.4.2015 1 Amniota
null
1 Lettre à Monsieur J.-C. Juncker, Président de la
A Debreceni Egyetem Egészségügyi Kar Jubileumi Lapja
ข ข่าว ฅ คน ปีที่ ๔ ฉบับที่ ๕ ประจำเดือนพฤษภาคม ๒๕๕๙
prezentace
KARTA MENU PROJEKT 1
Jak powstało ptasie pióro?
Strurovani_2015_vysledky
Télécharger Programme rentrée 2016
Wykład 2 - wbns.uksw.edu.pl
KatalogSTUR_2015
Pracovný list 12
Návod na použitie X3, X5
Yayın Değerlendirme/Book Reviews

![madarea2014_elso [Kompatibilitási mód].pdf](http://s1.readgur.com/store/data/000403430_1-cd91edee721270ead55e315fb3736b7b-260x520.png)