Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Lekce 2a (pdf)
TDL – přehled
Květen 2011
Rastlinné pletivá
Laboratorní cvičení č. 5 SPEKTROFOTOMETRICKÁ MĚŘENÍ ÚKOL
3. teoreticko-metodologické základy regionálnej geografie
chmielarz paweł - poland - Leśny Bank Genów Kostrzyca
Itinerár exkurzie - Katedra krajinnej ekológie
8. dermatogén (dermatogenum) (45/1D) — primârny meristém
5-mokřady ve světě
Anatomie a morfologie rostlin Rostlinné pletivo Orgány rostlin
Acta 89.indd - Výzkumný ústav Silva Taroucy pro krajinu a okrasné
Mikroskopická stavba dreva
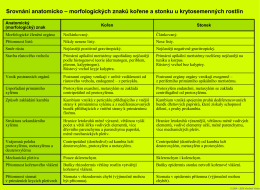
Srovnání anatomicko-morfologických znaků kořene a stonku
I rostliny mají hormony [pdf]
Lekce 5c
Seznam doporučené literatury
Metodika (PDF, 910 KB) - Výzkumný ústav Silva Taroucy pro krajinu
Kdy: středa 18. listopadu 2015od 18.30 do 20.00
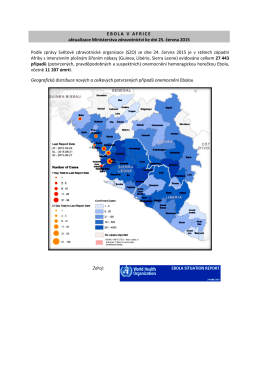
Ebola v Africe aktualizace MZČR ke dni 5. srpen 2015
zde
Przedmiar.pdf
dotazy rodičů












![I rostliny mají hormony [pdf]](http://s2.readgur.com/store/data/000152782_1-44a40b4f530386dabc9821997c460986-260x520.png)