Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
GENERAL BIOLOGY LABORATORY II
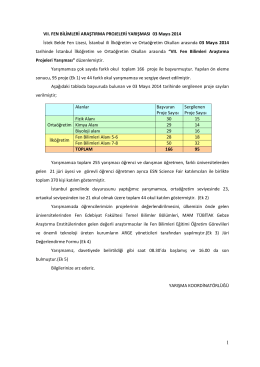
VII. FEN BİLİMLERİ ARAŞTIRMA PROJELERİ
"Vizyon ve Strateji ile Dönüşüm" isimli Sunumu
makaleyi indir
The Time of Gauss and Cauchy
DÜŞMAN
Toprak su kapasitesi - Ankara Üniversitesi Ziraat Fakültesi
KKTC Y AKIN DOGU UNiVERSiTESi MUHENDiSLiK F AKUL TESi
gida temel islemler I
03.06.15 / SELİN DAMAR (CANLI SAHNE) 04.06.15 / OTHERMAN
ÖZKUL "SÜPER KALiTE" CAMi HALISI ÖZELLiKLERi TOPLAM
kardiyoloji malzemeleri teknik şartnameleri
UÜ-SK BÜTÇE PLAN SATINALMA TEKNİK ŞARTNAME ÖRNEĞİ
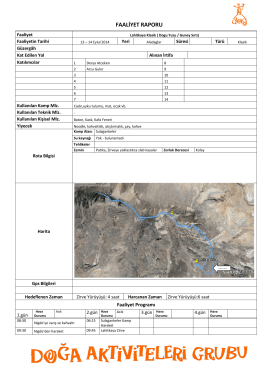
FAALİYET RAPORU
Kojenerasyon Için, Kompakt, Orta Boy, Kombine Çevrim
Program - KompEGE III
Odev-2 - Ahmet Sayar
ii I urvu L - Atelier KILICTURGAY
Kimya Bölümü Ders İçerikleri
KISA URUN BiLGiSt 1. BESERI TIBBI URUNUN ADI RECTODERM
3 1.ders - BOZOK ÜNİVERSİTESİ Personel Bilgi Sistemi
dersġn kodu - Ankara Üniversitesi Ziraat Fakültesi
EP DC 002 Kullanım Kılavuzu