Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Effects of the medial or basolateral amygdala upon social
Poster - WordPress.com
SODYUM SİLİKAT BAĞLAYICILI DÖKÜM KALIP/MAÇA KUM
4. Modulární příslušenství
Akademik Başarılarımız - Düzen Laboratuvarlar Grubu
dobrman 2012 - Dobrman Club ČR
İstatistikî İfadeyle... / Statistically Speaking
Oxytocín stimuluje remodeláciu tukového tkaniva potkana
panikové kovania katalóg Metalia
Şizofreni Genetiğinde Yeni Gelişmeler ve Bunların Klinik
yasal çerçeve ve alandan örnekler
zobacz artykuł
tıklayınız
Hangi hastalar Hepatit B virüs ilişkili karaciğer nakli sonrası yapılan
1. Name Surname: İbrahim Ömer Saatcioğlu 2
ZDE - Nebušice
Bilgisayar Terimler Sözlüğü
İçerik için tıklayınız
Mitral Kapak Replasmanı Yapılan Parkinsonlu
İndirmek İçin Tıklayınız - Süleyman Demirel Üniversitesi
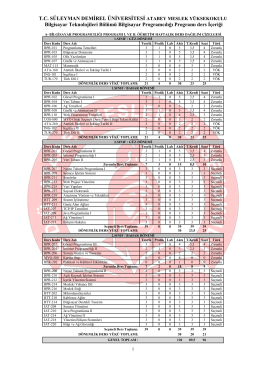
Bilgisayar Programcılığı - Atabey Meslek Yüksekokulu
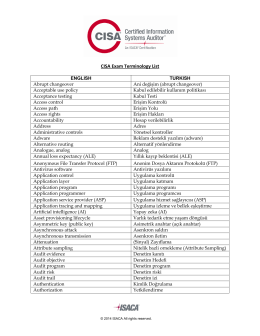
ISA-Exam-Terminology-List_xpr_Tur_1114
Ders 1